Microvesicles: Intercellular Vectors of Biological Messages
Cells communicate directly by cell-cell contact and indirectly via the release of mediators. But gaining a greater appreciation recently is the identification and characterization of intercellular communication through the secretion of microvesicles (MVs). MVs—small vesicles that comprise microparticles (MPs) and exosomes (1) —released from a wide variety of cells, can be considered micro-messengers. Whereas exosomes are released into the extracellular compartment by exocytosis, MPs are shed from the blebbing plasma membrane, and the composition and effects of both on target cells differ depending on the cell from which originate and the type of stimulus involved in their formation. MVs are obtained after several steps of centrifugation, whereby MPs are defined as the MVs obtained by centrifugation at < 100,000 × g, whereas exosomes are isolated by centrifugation at > 100,000 × g. Because the processes by which exosomes and MPs work are different, some researchers have claimed that such broad observations impede the defining and understanding of MV actions (2).
MPs are small vesicles, heterogeneous in size [(0.05–1 μm, a characteristic which is often used to distinguish MPs from exosomes (< 0.1 μm) and platelets (> 1 μm), respectively] and composition, with pro-coagulant and pro-inflammatory properties. Although all cell types can theoretically release MPs, the determination of the their origins has established that MPs can be released from the plasma membrane of circulating cells (such as platelets, erythrocytes, T and B cells, and monocytes); cells from the vascular wall (endothelial and smooth muscle cells); and tumor cells (3, 4). Even if the mechanisms governing MP formation are complex and not well understood, it is well appreciated that cell activation and apoptosis are the two main cellular processes that lead to their formation (Figure 1).
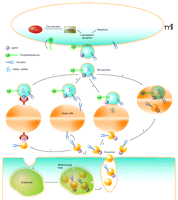
Once microparticles (top) and exosomes (bottom) are released, they can interact with target cells through four mechanisms: (1) ligand/receptor interaction, (2) protein transfer, (3) membrane fusion, or (4) internalization.
During cell activation by agonists or different stresses, MP formation is dependent on the influx of extracellular calcium and is associated with calpain activation, which is required for cytoskeleton disruption (5). In addition, blebbing and microvesiculation are often, but not always, preceded by the loss of membrane asymmetry resulting from local perturbation of the bilayer structure and the appearance of aminophospholipids such as phosphatidylserine, which is normally located in the inner leaflet, to the outer leaflet (6). The other main process of MP formation is apoptosis. Whereas MP blebbing occurs mainly during the early phases of apoptosis, apoptotic bodies are produced in the late stages of death process. Apoptotic bodies are larger than MPs and represent the compacted or condensed remnants of the shrinking apoptotic cells (7).
A better understanding of the mechanisms governing cell activation– and apoptosis-induced MP release would contribute to emergent therapeutic approaches that could decrease MP production in pathological conditions. The main limitation concerning studies related to MP formation, however, is that they are performed in vitro and very often with pharmacological inhibitors. It remains to be elucidated whether the same pathways are involved for the in vivo production of MPs.
An important parameter that determines the biological effects of MPs is their protein and lipid compositions, which may vary depending on the cell they originate from and the type of stimulus involved in their formation. MPs contain membrane, cytoplasmic, and nuclear constituents of their precursor cells, allowing for their characterization using antibodies directed against specific epitopes.
Exosomes are small, natural membrane vesicles released by a wide variety of cell types into the extracellular compartment by exocytosis. Exosomes were described initially during the secretion of MVs of endocytic origin by reticulocytes (8). Indeed, exosomes are formed within endosomes by invagination of the limiting membranes, resulting in the formation of multivesicular bodies. Subsequently, multivesicular bodies fuse with the plasma membrane and release exosomes into the extracellular environment (9). Secretion of exosomes can be spontaneous or induced depending on the cell type. Reticulocytes, T cells, mastocytes, and resting B cells secrete detectable levels of exosomes following the activation of a cell surface receptor. In contrast, dendritic cells, macrophages, and epithelial cells constitutively secrete exosomes in vitro, as do most tumor cells (10). The identification of MVs as exosomes is based on morphological and biochemical criteria. Thus, exosomes are obtained after high speed centrifugation (10); to confirm their presence and purity, immunoblotting analysis with antibody directed against exosomal markers like tetraspanins, heat shock protein (HSP)70, HSP90, or elongation factor-1α are needed. Also, electron microscopy or flow cytometry may be used for detection of exosomes, although there are claims that exosomes would be too small to be detected by flow cytometry.
Although exosomes do not contain any proteins from nuclear, mitochondrial, endoplasmic reticulum or Golgi apparatus, as a consequence of their endosomal origin, they are also selectively enriched in mRNA and miRNA, allowing genetic exchange between cells (11). Because MVs “hijack” the cytoplasm, capturing cytoplasm components and miRNA prior to release from the cell, the packaging of miRNAs in the cell may be random (12).
The mechanisms by which MPs and exosomes transfer biological information to recipient cells are not completely understood, but they may involve direct interaction with surface-expressed ligands or the transfer of surface receptors, proteins, mRNA, miRNA, and bioactive lipids. Moreover, they may serve as vehicles to transfer infectious particles (e.g., “Trojan horse” mechanisms) and to deliver intact organelles. Up to now, four mechanisms (ligand-receptor interaction, transfer, fusion, and internalization) by which information is transmitted by MVs have been reported (Figure 1, Table 1).
Different Mechanisms by which Microparticles and Exosomes Interact with Target Cells
Much research has demonstrated that ligands present on the surface of MPs and mediators released from MPs can directly stimulate receptors expressed on target cells. Thus, platelet MPs expressing P-selectin enhance leukocyte aggregation and accumulation by binding to P-selectin glycoprotein ligand-1 on the surface of leukocytes (13). Likewise, MPs harboring Sonic Hedgehog (Shh) promote megakaryocytic differentiation, nitric oxide (NO) production from endothelial cells, in vitro angiogenesis, and in vivo neovascularization by activating the Shh receptor. All of these effects are reversible with silencing of the Shh receptor (14–17). Additionally, MPs released from T cells can interact with smooth muscle cells through the Fas–Fas Ligand (FasL) pathway to evoke NF-κB activation (18).
Several studies point out that MPs may transfer receptors on the surface of target cells. Indeed, the release and intercellular trafficking of CD81+ MPs regulate the expression of CD81 surface receptors in lymphocytes (19). MPs might also participate in spreading HIV infection by transferring the CXCR4 co-receptor to CD4+/CXCR4− cells (20). Other instances of receptor transfer include arachidonic acid transport by platelet MPs to endothelial cells, leading to an increase in cyclooxygenase-2 (COX-2) and intercellular adhesion molecule 1 (ICAM-1) expression (21). Similarly, lipoxygenase 12 can be transferred from platelets to mast cells via MPs, leading to the production of the lipid mediator lipoxin A4, suggesting that platelet MPs pay a critical role as regulators of inflammation (22).
MPs can be also absorbed by fusion or by internalization (or engulfing). Fusion of MPs with their target cells leads to nonselective transfer of MP components (23). Tissue factor-rich MPs fuse with activated, P-selectin–expressing platelets, increasing the latter’s procoagulant activity. Concerning internalization, it has been shown that a mixture of MPs and exosomes released from endothelial progenitor cells are internalized in endothelial cells by interaction with α4 and β1 integrins. It is noteworthy that these MPs shuttle mRNAs that can activate angiogenesis in endothelial cells (24). Also, lymphocytic MPs interact with LDL receptors and can be internalized in tumor cells, decreasing tumor development (25).
Exosomes can bind to cells through receptor-ligand interactions. Dendritic cell–derived exosomes contain milk fat globule–epidermal growth factor 8 (MFG-E8) which can bind integrins expressed on dendritic cells or macrophages (26). Alternatively, exosomes can transfer information to recipient cells. Delta-like 4, a Notch protein family member whose expression is increased during angiogenesis, is present in exosomes and promotes capillary-like structure formation in vitro and in vivo by a mechanism implicating the transfer of Delta-like 4 into the endothelium, indicating that the Delta-like/Notch pathway does not require direct cell-cell contact to expand its signaling potential on angiogenesis (27). Furthermore, exosomes might attach or fuse with the target cell membrane, delivering exosomal surface proteins and cytoplasm to the recipient cell (28). Finally, exosomes may also be internalized by target cells by endocytosis or phagocytosis. Exosomes are internalized efficiently by phagocytes, by actin- and phosphatidylinositol 3-kinase-dependent mechanisms, without the need of caveolae, macropynocytosis, or clathrin-coated vesicles (29). Exosomes can also be internalized by immature dendritic cells for presentation to CD4 T cells (30).
A better understanding of intercellular transfer by exosomes may lead to the development of strategies that interfere with the release of pathogenic components by exosomes. Recently, a novel mechanism of genetic exchange between cells, referred to as “exosomal shuttle RNA,” by a transfer of mRNAs and miRNAs mediated by exosomes, has been proposed (11), opening the possibility of using exosomes for gene therapy. Also, exosomes secreted by dendritic cells could modulate immune responses directly by exposing MHC and T-cell costimulatory molecules and indirectly by carrying internal components to surrounding cells (10, 31).
Among the different messages transferred, MVs are able to modify function of cells from the cardiovascular system, immune system, and tumor cells. The transfer of information between MVs and cells is independent of the number of MVs but is dependent on the different composition and origin of the MVs (32, 33). Owing to the procoagulant ability of tissue factor and of phosphatidylserine that MVs carry, elevated levels of MPs participate in the development and maintenance of prothrombotic status in atherosclerosis (34), pre-eclampsia (35), and hematological diseases (36), as well as in cancer-associated thrombosis (37).
Circulating MPs decrease NO production by either reducing the activity of endothelial NO-synthase or by decreasing its bioavailability. Thus, MPs from patients with myocardial infarction, diabetes, or pre-eclampsia induce endothelial dysfunction by impairing the endothelial NO transduction pathway (38–40). In patients with metabolic syndrome, circulating MPs evoke endothelial dysfunction by decreasing endothelial NO-synthase activity and NO production by increasing protein nitration in endothelial cells (41) and by enhancing plasmatic oxidative stress markers (42). Interestingly, patients with obstructive sleep apnea (OSA) display the same levels of circulating MPs as observed in healthy subjects, but levels of MPs from granulocytes and activated leukocytes (CD62L+) are higher in OSA patients. In addition, levels of CD62L+ MPs correlate with endothelial-NO dysfunction that may initiate atherogenic processes in patients with OSA (32). In a model of rat pulmonary arterial hypertension, circulating MPs from hypoxic rats exhibit reduced NO bioavailability owing to decreased endothelial NO-synthase activity and enhanced oxidative stress in pulmonary endothelial cells (33). Altogether these data underscore the deleterious effects of circulating MPs on the cardiovascular system, and in particular, on the endothelial cells.
Finally, one key feature of the effects of MVs in the cardiovascular system is their ability to modulate the angiogenic program of both endothelial mature and progenitor cells. Indeed, proteins such as peroxisome proliferator–activated receptor–alpha (PPARα), carried by MPs, seem essential to promote the pro-angiogenic reprogramming of endothelial progenitor cells through Akt- and NF-κB–mediated pathways (43). Isolated MPs from vitreous fluid from patients with diabetic retinopathy stimulated endothelial cell proliferation and increased new vessel formation in mice, indicating that MPs could contribute to disease progression through their ability to promote angiogenesis (44). MPs are able to amplify the initial endothelial dysfunction and accelerate the progression of atherosclerotic lesions by promoting intraplaque neovascularization and thrombogenicity (45).
Endothelial exosomes bearing metalloproteinases promote the formation of a tubule-like structure in human endothelial cells, suggesting that exosomes may regulate proteolytic activity vital to angiogenic steps involving migration of endothelial cells (46). Altogether, these data highlight the potential effects of circulating MVs on new vessel generation and may constitute a new therapeutic target against the progression of these diseases.
Immune cells are activated during many pathological situations and subsequently generate MVs. MPs appear to serve as important mediators to promote inflammation whereas exosomes most often drive immune responses and autoreactivity. Indeed, through the generation of MPs, activated T cells may facilitate activation of mast cells that are not located in close physical proximity at the inflammatory sites (47). In addition, platelet-derived MPs may play an important role by acting as vectors for the CD154 (CD40L) signal. CD154 is a member of the TNF family expressed by activated CD4+, CD8+, and γδ-T cells, B cells, and platelets. Among the effects of CD154, the CD154 signal plays a crucial role in the activation of T cell–dependent humoral immune responses (48). In particular, by concentrating the CD154 activity and carrying it away from the site of platelet activation and aggregation, platelet-derived MPs stimulate antigen-specific production by cooperating with responses elicited by CD4+ T cells (49).
Depending on their origin, exosomes can serve as both immunostimulatory and immunoregulatory entities (50). As described above, T and B cells as well as dendritic cells release exosomes. Thus, in vitro studies have shown that exosomes released from B cells can directly activate CD4+ T cells (51). Also, dendritic cell-derived exosomes initiate specific cytotoxic T lymphocyte response in vivo and suppress growth of established murine tumors in a T cell-dependent manner (52). In the same way, tumor-derived exosomes transfer tumor antigens to dendritic cells, which induce potent CD8+ T cell–dependent antitumor responses (53). Subsequently, numerous studies have reported the ability of exosomes to act as potential relevant elements for immuno-interventions by amplifying the generation of donor-reactive T cells following transplantation (54) or by suppressing antigen-specific responses through a Fas/FasL-dependent mechanism in the treatment of autoimmune diseases (55).
The successful quantification and identification of several proteins carried by exosomes allows for the suggestion that exosomes may be useful as prognostic, diagnostic, or detection markers in cancers. For instance, claudin-4 can be shed from ovarian cancer cells and detected in exosomes from plasma from ovarian cancer patients (56). The number of exosomes expressing CD63 [a tetraspanin (a cell surface–localized membrane protein) involved in several cellular process including cell activation, adhesion, differentiation, and tumour invasion (57)] and caveolin-1 in the plasma of melanoma patients is significantly increased when compared to those in healthy donors (58). Also, the cytochrome P450 CYP17A1, a target for total androgen blockade in advanced prostate cancer patients, is expressed in plasma exosomes for these patients (59), whereas the presence of two known prostate cancer biomarkers, PCA-3 and TMPRSS2:ERG, have been found in exosomes isolated from urine of patients (60), suggesting that exosomes may be used as a novel approach in detection and prostate tumor surveillance. Also, exosomes from glioblastoma tumor cells containing mRNA, miRNA, and angiogenic proteins, such as VEGF, can be detected in serum from glioblastoma patients. Thus, exosomes promote angiogenesis favoring cancer growth, yet they can provide valuable information regarding the diagnosis and choice of appropriate therapy (61). Cancer specific miRNA isolated from circulating tumor-derived exosomes from patients with ovarian cancer constitutes a new biomarker of this pathology (62).
Most interestingly, exosomes can represent a nexus between the immune system and tumor environment. Isolated exosomes from the blood of patients with colorectal cancer (63) contain not only proteins typical of exosomes (CD63) but also FasL and TNF-alpha-related apoptosis-inducing ligand (TRAIL), which mediate their pro-apoptotic effects on CD8+ T cells (63). This mechanism of immunosuppression has potential implications as a prognostic factor and could be targeted for the development of new antitumor therapies in colorectal cancer patients. Also, exosomes present in blood and ascites from patients with ovarian carcinoma promote differentiation and proliferation of regulatory T cells (Treg) as well as up-regulation of Treg-mediated suppression that may potentially contribute to tumor escape (64). Unlike helper T cells, which orchestrate antibody production by B cells, and cytotoxic T cells, which kill virally infected or abnormal cells, Treg act to suppress the immune response of a large number of distinct target cell types (65). Furthermore, these exosomes injected into tumor-bearing mice resulted in augmented tumor growth (66). Altogether, exosomes act as vectors for the transfer of immunological information at distance and can be considered as novel biomarkers and effectors.
In summary, MVs display two faces, as did the Roman god Janus: 1) they constitute potential biomarkers of activity disease, and 2) they can be considered as promising therapeutic targets, considering their ability to convey biological information.
Acknowledgments
This work was partially supported by Agence Nationale pour la Recherche- 07-PHYSIO-010-01 and Fondation de France (n° 2007001918 ).
- Copyright © 2011
References
Simon Tual-Chalot is a PhD student at the Institut National de la Santé et de la Recherche Médicale (INSERM) U694 in the University of Angers, France. His research aims are to determine the vascular implication of microparticles in pneumological disorders, especially obstructive sleep apnea.

Daniela Leonetti, PhD, is currently a Research Engineer at INSERM U694, Mitochondria: Regulation and Pathology, in the University of Angers. Her PhD studies, under the supervision of Ramaroson Andriantsitohaina, focused on the role of membrane microparticles in the pathophysiology of Crohn’s disease.

Ramaroson Andriantsitohaina, PhD, is Director of Research at the Institut National de la Santé et de la Recherche Médicale (INSERM) at INSERM U694 in the University of Angers, France. His main interest focuses on translational research depicting the role of oxidative stress in metabolic and inflammatory diseases with special interest for microparticles and plant polyphenols. Send correspondence to RA: E-mail: ramaroson.andriantsitohaina{at}univ-angers.fr; fax +33 2 44 68 85 88.

M. Carmen Martinez, PhD, is Director of Research at the Centre National de la Recherche Scientifique at INSERM U694 in the University of Angers, France. Her research focuses on the cellular and molecular effects of microparticles on endothelial function and angiogenesis.


