A Bacterial Sensory System that Activates Resistance to Innate Immune Defenses: Potential Targets for Antimicrobial Therapeutics
Over the course of thousands of years involving countless interactions with their hosts, successful pathogens have evolved mechanisms to avoid or subvert host defenses, and in the case of Salmonella enterica, a number of virulence factors act in concert to establish and promote bacterial infection. One such factor is the PhoP-PhoQ two component regulatory system. PhoQ is an inner membrane sensor-kinase composed of a periplasmic sensor domain and a cytoplasmic kinase domain that phosphorylates its cognate transcription factor, PhoP, upon perception of specific environmental signals. Like other members of the response regulator family, phos-pho-PhoP binds DNA and activates or represses its target genes. The PhoP-PhoQ system responds to changes in the concentration of divalent cations––it is repressed by millimolar and activated by micromolar concentrations of magnesium (1)––to activate or repress transcription of a wide variety of bacterial genes. A subset of the genes activated by PhoP are involved in the remodeling of the outer membrane, which serves to stabilize the bacterial envelope in the absence of divalent cations and enables bacterial growth in medium lacking magnesium. The ability of Salmonella to grow in the presence of limiting concentrations of magnesium is abrogated in the absence of PhoP or PhoQ. Studies on the mechanism of the low magnesium response suggested that magnesium binds directly to the PhoQ periplasmic domain, leading to a conformational change and consequent gene regulation (1).
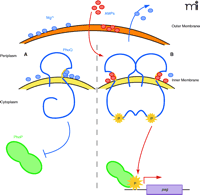
The interaction of magnesium and AMPs with theSalmonella entericaPhoP-PhoQ two-component regulatory system. PhoP-PhoQ is required for survival in the presence of low magnesium, antimicrobial peptides [(AMPs), i.e., PB, PG-1, CRAMP, and LL-37], cultured macrophages, and within animal hosts. Both magnesium ions (blue ovals) and AMPs (red ovals) are thought to bind an acidic pocket in the periplasmic domain of PhoQ at the inner membrane interface (yellow patch). Millimolar concentrations of magnesium repress the PhoP-PhoQ system (A), whereas sublethal concentrations of AMPs displace magnesium and activate this system (B). Administering AMPs to bacteria leads to loss of Mg2+ ions from the bacterial membrane as well as from PhoQ itself. Activation of PhoQ is the result of a conformational change in the PhoQ protein that promotes dimerization and autophosphorylation, with subsequent phosphotransfer to the DNA response regulator, PhoP. Phospho-PhoP activates transcription of a number of genes (pag) that promote survival of Salmonella enterica in the host.
More recently, studies have shown that the transcription of PhoP-activated genes is activated not only by low magnesium, but by the presence of sublethal concentrations of antimicrobial peptides (AMPs) (2, 3). These peptides are key components of the host innate immune defense response, functioning at mucosal surfaces as part of the barrier that prevents pathogen colonization of host tissues, and within phagocytic cells of the immune system to kill engulfed or invasive microorganisms (4, 5). AMPs are small cationic molecules that can form pores in microbial membranes and are produced by virtually all living organisms. Because AMPs target conserved aspects of microbial physiology that enable specific killing of microbes with minimal toxicity to the host, they are potentially very attractive as a basis for antimicrobial therapeutics (5); however, many microbial pathogens possess mechanisms of resistance to AMPs. In particular, just as the Salmonella PhoP-PhoQ system enables bacterial growth in low magnesium environments, it also promotes bacterial resistance to a wide variety of structurally unrelated AMPs [notably, polymyxin B, magainin, protegrin-1, and cathelicidin-related AMP (CRAMP)], which have markedly greater activity against phoP or phoQ mutant Salmonella than to corresponding wild-type bacteria (2, 6–8).
PhoP-PhoQ-mediated outer membrane modifications that occur in the absence of magnesium during in vitro culture conditions have also been observed to occur within mammalian macrophage cells in tissue culture (9). These modifications are presumably required for successful bacterial growth within the macrophage vacuole as phoP mutant bacteria are deficient not only for growth in low magnesium medium or in the presence of AMPs, but also within macrophages and consequently in mouse and human infections as well (10, 11). Elucidating the mechanism by which the PhoP-PhoQ system promotes AMP resistance in bacteria has understandably been a question receiving a great deal of recent scientific interest. Interfering with the activation of PhoP-PhoQ in vivo would render Salmonella avirulent; therefore, PhoP-PhoQ is a prime target for therapeutic intervention. In addition, phoPQ mutant strains have received considerable attention as live-attenuated vaccines (12).
A recent study by Bader et al. (13) on the precise mechanism of how AMP are sensed by the PhoP-PhoQ system has suggested that, in a manner similar to that of divalent cations, AMPs bind directly to the periplasmic domain of PhoQ; however, this binding activates rather than represses the PhoP-PhoQ system, leading to transcription of PhoP-activated genes. As a consequence, exposing bacteria to sublethal concentrations of AMPs leads to a typical adaptation stress response, with the subsequent ability of bacteria to survive increased concentrations of AMPs. It is hypothesized in this work that magnesium and AMPs bind to the same acidic pocket of the periplasmic domain of PhoQ. Structural data demonstrates that this anionic patch faces the inner membrane, which puts it in a good position to interact with the cationic, amphipathic peptides that tend to lie on membrane surfaces. It is unclear from this work, however, if the subsequent activation of PhoQ is the result of local displacement of magnesium from the acidic pocket or, as proposed by Bader et al., that the binding of AMPs to the periplasmic domain of PhoQ directly activates its kinase functions. In either case, it may be possible to devise therapeutic AMPs that retain their pore-forming activity against the bacterial membrane, yet either do not bind to PhoQ, or bind but do not induce a structural change in PhoQ (much like in the case of magnesium).
It is intriguing that AMPs that are unrelated in either primary sequence or tertiary structure are all capable of activating PhoQ, suggesting either that the periplasmic domain has a fair amount of plasticity in terms of molecules that it can bind (merely electrostatic interactions between the anionic periplasmic patch and the cationic peptides), or that there is an as-yet-unidentified common feature to the peptides that can activate PhoQ. It would thus be of great interest to examine the structural basis for the interaction of various peptides with PhoQ, and, furthermore, to characterize bacterial responses to peptides that have antimicrobial activity but that fail to activate PhoQ.
Bader et al. further showed that a chimeric PhoQ protein, in which the endogenous periplasmic domain had been replaced with that of the PhoQ homolog of Pseudomonas aeruginosa, retained its ability to respond to low magnesium but failed to signal in response to AMPs. This finding raises a number of interesting questions. In particular, because other studies have shown that the PhoP-PhoQ system of Pseudomonas is also involved in AMP resistance, it suggests that Pseudomonas and Salmonella may sense AMPs via different mechanisms. This possibility is strengthened by the observation that phoQ mutants of Pseudomonas show constitutive activation of PhoP-regulated genes (14). This is in surprising contrast to Salmonella and E. coli, in which phoQ mutants are unable to respond to magnesium or AMPs. This is an important distinction from the standpoint of developing antimicrobial peptide-based therapeutics because such therapeutics may have to be tailored for specific classes of microbes that are distinguished not by whether they are Gram-negative or Gram-positive, but rather by the mechanism through which they sense AMPs. An additional caveat to the development of AMPs as therapeutics is the possibility that pathogenic microbes will develop wide-scale resistance, as has occurred with previous generations of conventional antibiotics. AMPs serve as attractive therapeutic candidates because resistance to AMPs does not confer cross-resistance to conventional antibiotics and because AMP resistance is generally difficult to acquire. On the other hand, because AMPs comprise such an important part of our own innate immune response, the wide-scale use of AMP antibiotics is something that should be approached with extreme caution. Recent studies demonstrate that microbes can evolve resistance to AMPs (15). Thus, rather than being the magic bullet for solving the problem of antimicrobial resistance, AMP therapy could potentially enable microbes to bypass or overcome the innate immune response of an immunocompetent host.
- © American Society for Pharmacology and Experimental Theraputics 2005
References

John Gunn, PhD, is an associate professor in the Department of Molecular Virology, Immunology, and Medical Genetics at The Ohio State University with an interest in the molecular mechanisms used by Salmonella spp. to survive harsh conditions (primarily antimicrobial peptides) it encounters within the human host at mucosal surfaces and within the macrophage phagosome. He has been involved in many of the seminal studies that established the importance of PhoP-PhoQ and downstream regulatory systems in virulence, antimicrobial peptide resistance, and outer membrane modification. E-mail: gunn.43{at}osu.edu; fax 614-292-5495.

Igor Brodsky, PhD, is a postdoctoral fellow at the Yale University School of Medicine in the laboratory of Dr. Ruslan Medzhitov. He received his PhD from Stanford University in 2003 where he trained in the laboratory of Dr. Stanley Falkow. His PhD thesis work focused on the role of a particular PhoP-regulated gene in resistance to antimicrobial peptides. His current post-doctoral studies are on the interactions of the host innate immune response with Yersinia pseudotuberculosis.

