Is a Step Backwards in S-Phase-Targeted Chemotherapy a Step Forward?
It is a difficult task to ensure that the approximately three billion bases in the genome are undamaged and correctly aligned. It is even tougher when that number doubles to six billion base pairs during DNA replication. Fortunately, cells possess an intricate network of checkpoints that ensure the faithful reproduction and ultimately integrity of the genome. Although there has been considerable progress in our understanding of the cell cycle checkpoints, two fundamental questions remain: what are the signals that trigger the checkpoint response, and how does the cell recognize this signal? Using orthogonal approaches, two recent studies (1, 2) have clarified what does and does not trigger a checkpoint response and may have identified a new cancer chemotherapeutic target in an established cellular process.
During the DNA synthesis (S) phase of the cell cycle, a replicating cell must duplicate the genome exactly so that an identical copy can be passed to each daughter cell during mitosis. Replication of DNA is commonly portioned into three steps: origin licensing, replication initiation, and replication elongation. During phase, the proteins making up the orilate mitosis and early G1 gin replication complex (ORC), Cdc6, and the helicase proteins Mcm2-7 associate with a number of specific DNA sequences in a process called origin licensing. Once S phase has formally begun, origin firing occurs, in which the kinases Cdc7 (associated with its regulatory subunit Dbf4) and Cdk2 phosphorylate several proteins associated with replication initiation, including Cdc45 and the Mcm helicase complex. Phosphorylation of these proteins triggers the binding of DNA polymerase to the origin of replication, and the Mcm helicase begins to unwind the double helix ahead of replication to form the replication fork. This process leads to the elongation step, during which the replication fork becomes fully formed and the DNA replication complex moves along the DNA until it is joined at another replicon.
Accurate replication of the genome requires a tremendous amount of energy and coordination; improper spatial or temporal control of DNA synthesis can have catastrophic consequences for the genome. Thus, cells have evolved mechanisms to sense problems with the replication and segregation of DNA into daughter cells, and these are known as cell cycle or DNA damage “checkpoints.” These checkpoints are mediated by a hierarchy of signaling proteins officiated by DNA-sensing enzymes, which identify aberrant DNA structures such as single-strand and double-strand DNA breaks, missing or modified DNA bases, and stalled, malformed, or collapsed replication forks. The damage is transmitted by complex pathways that influence cell cycle progression, DNA repair, and cell survival.
During elongation, collapse of the replication fork or collision with DNA adducts can lead to DNA strand breaks, which, if left unrepaired, can trigger cell death. Some of the most cytotoxic anti-cancer agents to date target the elongation step of DNA replication by inducing either direct or indirect DNA damage. Generally there are three modes of replication fork disruption: inhibition of DNA polymerase activity; restriction of available intracellular deoxynucleotide pools; and physical impedance of the replication fork via adduct formation or DNA strand cross-linking (Figure 1A⇓). The checkpoint response had been thought to be initiated only in the presence of actual DNA lesions, such as those described above. Recent studies by Montagnoli et al. (1) and Soutoglou and Misteli (2), however, indicate that actual DNA damage may be unnecessary and that targeting the S phase of the cell cycle without the overt generation of DNA damage or checkpoint activation is possible, if the focus is shifted from elongation to initiation.
DNA damage sensors such as the Mre11-Rad50-Nbs1 (MRN) complex, which recognizes DNA double-strand breaks, are well known, but the communication networks that transmit the message of damaged or aberrantly-structured DNA to the checkpoint effector proteins are not fully established. Now, the absolute requirement of DNA damage in the DNA damage response has been challenged. Soutoglou and Misteli (2) found that tethering of the MRN complex proteins MRE11 and NBS1 (as fusion proteins containing the lac operator) to the LacO site in human DNA can activate the DNA damage checkpoint response even in the absence of bona fide DNA damage. Based on their work, it appears that prolonged localization of at least some DNA repair factors to undamaged chromatin can trigger a checkpoint response analogous to that produced following exposure to DNA damaging agents. This unexpected observation suggests that the actual “sensing” of DNA damage or aberrant DNA structures may not be required for the activity of the checkpoint pathway and draws into question our current models of checkpoint activation.
What is the actual trigger that promotes downstream signaling? The observations of Soutoglou and Misteli (2) suggest that damage sensing factor “dwell-time” at a particular site in DNA may be a determinant in the activation of the checkpoint response. This stimulates one to consider applying notions of receptor occupancy that, heretofore, were restricted to plasma membrane receptors to DNA itself (3). These surprising experimental results also raise the question of what prevents these proteins from “loitering” at specific sites on chromatin under unstressed conditions.
If temporal and location constraints of damage-sensing proteins and not binding per se to aberrant DNA structures is truly responsible for pathway activation, then one wonders how do the sensors accumulate at sites of DNA damage? Several possibilities exist. First, and maybe the most likely, is that the MRN complex or specific components of MRN act as DNA scanners, whose progression along the length of DNA strands is impeded by strand breaks and collapsed or malformed replication forks. This otherwise logical hypothesis is contradicted by the notion that DNA exists primarily in complex with histones and nonhistone regulatory proteins; is displacement of these proteins required for MRN activity? Further analysis is required for a full understanding of the role of individual DNA damage sensing and repair enzymes and their relative importance in the DNA damage response cascade once bound to DNA. Harnessing agents that modulate the binding of DNA damage response molecules to DNA may serve as platforms for lead compounds, providing entirely new classes of anticancer agents. Whether protein “tethering” to chromatin alters cell viability remains to be examined.
Recent studies by Montagnoli et al. (1) provide a clever pharmacological approach to inhibit DNA replication in the absence of traditional checkpoint activation. They identified PHA-767491, a potent, selective inhibitor of the kinase Cdc7, by screening >450,000 compounds in a biochemical kinase assay. Treatment of cancer cells with PHA-767491 inhibited Cdc7 activity in vitro, inhibited phosphorylation of Cdc7-dependent sites on its physiological target Mcm2 in cells, and caused accumulation of cells at the G1-S border and also in G2-M phase of the cell cycle. Moreover, this Cdc7 inhibitor exhibited efficacy against a variety of human tumor cell lines, including those that are resistant to conventional chemotherapeutics targeting the elongation phase of DNA synthesis, such as gemcitabine and 5-fluorouracil. PHA-767491 caused impressive growth inhibition of tumor xeno-grafts in mice and carcinogen-induced tumors in rats without significant toxicity.
Because Cdc7’s kinase activity is necessary for the initiation of DNA replication, a chemical inhibitor of this kinase activity would be expected to halt progression into S phase. The authors used a technique called “DNA combing” in which newly synthesized DNA is fluorescently labeled and stretched out on glass slides for visualization to examine the mechanism by which PHA-767491 caused cell cycle arrest. They found PHA-767491 inhibited the firing of replication origins but did not significantly affect the progression of replication forks from origins fired prior to drug treatment as is the case for conventional S phase–acting chemotherapeutics, indicating PHA-767491 exerts its effects at the replication initiation stage. To date, this is the first report of a chemical agent that acts during the initiation phase of DNA replication, thus preventing DNA synthesis before it has begun.
Intriguingly, inhibition of replication by PHA-767491 results in cell cycle arrest and induction of apoptosis but no initiation of the DNA damage response; presumably this is because the replication fork associates with ATR [ataxia-telangiectasia-mutated (ATM) and Rad3-related protein], which is necessary for initiating checkpoint signaling from the replication fork. As the fork has not yet formed in cells halted at the initiation phase, no checkpoint signaling can occur. This pharmacological blockade of S-phase progression without activating the checkpoint is congruent with other reports in which knockdown of Cdc7 with siRNA caused S-phase arrest but failed to elicit a checkpoint response (4).
The observation that PHA-767491 potently induces cancer-cell death raises several questions. First, this compound exhibits impressive selectivity, potently inactivating Cdk9 and Cdc7. It remains unclear, however, whether the inhibition of Cdk9 is necessary for PHA-767491–mediated apoptosis: genetic inhibition of Cdk9 has quite varying effects on cell growth and viability [discussed in (1)], suggesting Cdk9 inhibition may not be an important contributor to the pharmacological actions of PHA-767491. In support of a Cdc7-dependent mechanism, suppression of Cdc7 either by conditional knockout in mouse embryonic stem cells or by siRNA in p53-depleted normal fibroblasts and p53-deficient HeLa cervical carcinoma cells induced an abortive S phase and subsequent death by p53-independent apoptosis or mitotic catastrophe (4, 5). The cell death induced by PHA-767491 appears to be at least partially dependent upon defective p53 signaling, as p53-positive normal cells do not display characteristics of apoptosis upon treatment but become sensitized after exposure to p53 siRNA. This finding is in agreement with previous reports of siRNA-mediated suppression of Cdc7 that elicit apoptosis in the absence of p53 (4). Intriguingly, PHA-767491 kills cancer cells independent of p53 status, indicating that in cancer cells the inhibitor may exert its effects via a currently unknown process that includes Cdc7 inhibition. Left unanswered is the mechanism of G2-M arrest. Cdc7 has roles both in mitosis and in cytokinesis in yeast (6, 7), and a recent report suggests that Cdc7, when associated with an alternative regulatory subunit named Drf1, may facilitate progression through M-phase (8). Inhibition of Drf1 using siRNA results in the attenuation of cellular proliferation with a marked in increase in cells in the G2-M phase of the cell cycle (8).
Do these two articles portent a new wave of cancer therapeutics? The notion that checkpoints can be exploited for cancer therapy is not new. These two studies, however, might stimulate novel approaches that capitalize on the idea that a functional checkpoint can be initiated in the absence of DNA damage and that alternate steps in replication can result in selective cancer cell killing without checkpoint activation. The process of licensing origins has not to our knowledge been exploited therapeutically, and many additional targets exist in the initiation complex. The multiple regulatory subunits of Cdc7 provide the opportunity to selectively disrupt specific Cdc7-dependent cellular processes if Cdc7 does, in fact, regulate cellular processes aside from DNA replication.
These studies expose weaknesses in the regulation of DNA replication and sensing of DNA damage that might be exploited therapeutically. PHA-767491 represents one promising new agent that targets Cdc7 with antitumor activity and limited toxicity in animal models. Moreover, for supporters of high throughput screening of large chemical libraries, it affirms the validity of in vitro kinase assay screen in the search for new antineoplastic agents. The notion that DNA damage is not required for the activation of the checkpoint response also presents new avenues for potential therapeutic intervention and the discovery of agents capable of modulating the temporal and spatial MRN protein binding to DNA should assist in clarifying the process of checkpoint activation.
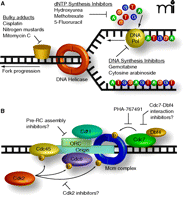
Current and hypothetical targets for antineoplastic therapy targeting DNA synthesis. A. FDA-approved cancer drugs that target S-phase cells disrupt the replication elongation stage, during which the replication fork is already formed. Collision of the replication fork with DNA adducts can induce DNA strand breaks, which activate the S-phase checkpoint. Similarly, the progression speed of the fork is monitored by the transducer kinase ATR. If the fork is destabilized or slowed significantly, ATR mediates checkpoint activation. Thus, the current chemotherapeutics activate a replication fork-dependent checkpoint. B. PHA-767491 targets the kinase Cdc7, which is required for the replication initiation phase. In the initiation phase, the replication fork is not yet formed, thus, interfering with this step of DNA synthesis does not elicit a checkpoint response. The efficacy of PHA-767-491 in multiple cellular and animal models, combined with its apparent selectivity for cancerous cells suggest that targeting the replication initiation machinery may be a novel and effective method for chemotherapy.
Footnotes
-
Note added in proof: During the preparation of this article, the authors became aware of an additional study describing checkpoint activation in the absence of DNA damage in yeast (9).
- © American Society for Pharmacology and Experimental Theraputics 2008
References

Pierre E. Queiroz de Oliveira, MS, Graduated in 2002 from the University of Bath with a Masters in Pharmacology and is currently a pharmacology graduate student in John S.Lazo’s laboratory. He is exploring the role of oxygen deprivation in the regulation of cell cycle phosphatases.

Robert J. Tomko, Jr., BS, is a pharmacology graduate student in John S. Lazo’s laboratory at the University of Pittsburgh. He is currently studying the role of oxidative and nitrosative post-translational modifications in the regulation of tyrosine phosphatase activity.

John S. Lazo, PhD, is the Allegheny Foundation Professor of Pharmacology and the Director of the University of Pittsburgh Drug Discovery Institute. After graduating with a bachelor degree in Chemistry from the Johns Hopkins University, he completed his PhD in Pharmacology with Raymond Ruddon at the University of Michigan. He joined Alan Sartorelli as a postdoctoral fellow and remained on the faculty in the Department of Pharmacology at Yale University until 1987, when he became Chairman of Pharmacology at the University of Pittsburgh. His laboratory is currently interested in the biological role of protein phosphatases in normal and malignant cells and in the pharmacological actions of novel natural products. E-mail: lazo{at}pitt.edu; fax 412-648-9009.

