γ-Secretase: A Catalyst of Alzheimer Disease and Signal Transduction
Abstract
γ-Secretase is a membrane-bound protease that cleaves within the transmembrane region of the amyloid precursor protein to generate the C termini of both major peptides that constitute the amyloid plaques found in Alzheimer disease (AD). Recent biochemical studies provide compelling evidence that presenilins, membrane-spanning proteins associated with early-onset familial AD, are novel aspartyl proteased that function within a macromolecular complex to mediate γ-secretase activity.These new insights underscore the role of the presenilins in regulated intramembrane proteolysis and could possibly lead to the inhibition of -secretase activity as a therapy for AD.

Proteolytic processing of the amyloid precursor protein (APP) is fundamental to the plaques that develop in brain tissue afflicted
by Alzheimer disease (see above). The processing of APP, not only in Alzheimer disease, but also in normal physiology, is
thus a major focus of research.
INTRODUCTION
In view of the world's rising elderly population, Alzheimer disease (AD), an age-related neurodegenerative disorder, threatens to precipitate a public health crisis. Although there has been substantial progress in the last decade in understanding the molecular neuropathogenesis of AD, there is no treatment that significantly affects onset or progression of the disease. Histopathologically, AD is characterized by the presence of extracellular amyloid plaques and intracellular neurofibrillary tangles in the brain. In accordance with the amyloid cascade hypothesis (1), β-amyloid peptide (Aβ), the major constituent of amyloid plaques, is believed to play a central role in the neuropathology of AD (Figure 1⇓). Aβ generally exists in two forms, either 40 or 42 amino acids in length, that differ at their C termini. Of the two forms, Aβ42 is more prone to aggregation (2).
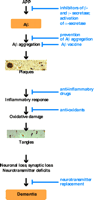
Pathological pathways of AD and potential sites of therapeutic intervention. It is hypothesized that Aβ plays the most important role in the initiation of the neuropathogenesis of AD. Other factors also contribute to the onset and progress of the disease. Amyloid plaques and neurofibrillary tangles are hallmarks of the disease. Therapies that might reduce Aβ production, mitigate plaque formation, or otherwise interfere with the progression of AD are indicated in blue.
PROCESSING OF THE AMYLOID PRECURSOR PROTEIN
Aβ is derived from the processing of the amyloid precursor protein (APP) by two proteases, β-secretase and γ-secretase, that generate the N and C termini of Aβ, respectively (Figure 2⇓). Alternatively, APP can be processed by α-secretase, which precludes the formation of Aβ. Because the secretases are involved in the earliest steps of the pathogenic pathway of AD, they are prime targets for drug development.
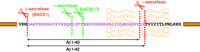
The amyloid precursor protein (APP) can undergo multiple cleavage events. Aβ is derived from APP by β- and γ-secretase–mediated cleavage. Like α-secretase, BACE2 probably cleaves within the Aβ sequence and prevents formation of the Aβ40 and Aβ42 peptides that lead to amyloid plaques.
Although cleavage of APP by α-secretase is a major proteolytic pathway, the exact identity of α-secretase has not been confirmed. Preliminary data suggest that TACE (for TNF-α converting enzyme), a protease that belongs to the ADAM family (for a disintegrin and metalloprotease), may have α-secretase activity (3). Despite this lead, α-secretase may not be the most attractive target for pharmaceutical intervention of AD, because the therapeutic upregulation of α-secretase poses greater logistic challenges than does the chemical inhibition of the β- and γ-secretases.
After more than a decade of searching for the identities of APP processing proteases, the enzyme that manifests β-secretase activity was recently discovered (4–,7). BACE1 (for β-site APP-cleaving enzyme; also known as Asp2, or memapsin 2) is a type-one transmembrane aspartyl protease with the essential characteristics expected of β-secretase: 1) BACE1 overexpression leads to increased levels of β-secretase products; 2) BACE1 antisense probes decrease β-secretase products; 3) BACE1 is expressed in the brain and co-localizes at the subcellular level with APP; and 4) purified endogenous or recombinant BACE1 cleaves APP and synthetic peptide substrates at the scissile bond recognized by β-secretase. The crystal structure of BACE1 in association with a hydroxyethylene transition-state inhibitor has been solved (8) and reveals a bilobal structure that resembles the conserved general folding of aspartyl proteases. The active site of BACE1, however, is more open and less hydrophobic than those of other human aspartyl proteases.
Knockout technology has been used to validate the function of BACE1 in the production of Aβ (9, ,10). BACE1 knockout mice (BACE1-/-) appear normal with respect to gross anatomy, tissue histology, hematology, and clinical chemistry. Nevertheless, secretion of Aβ is abolished in cultures of BACE1-deficient embryonic cortical neurons. When BACE1-/- mice are crossed with transgenic mice that overexpress APP, neither Aβ nor β-CTF (β-secretase–cleaved APP C-terminal fragment) can be detected in the brains of the progeny mice. These studies establish that BACE1 is the principle β-secretase in vivo and suggest that BACE1 inhibitors may be free of mechanism-based toxicity in the treatment of AD.
Another aspartyl protease, BACE2 (Asp1, memapsin 1), a BACE1 homolog, has also been discovered. Although its tissue expression profile does not fit the criteria of β-secretase, BACE2 nevertheless recapitulates the in vitro substrate specificity of β-secretase. Additionally, BACE2 cleaves within Aβ; BACE2 may thus have a function similar to α-secretase in the prevention of Aβ production (Figure 2⇑) (11, ,12).
γ-SECRETASE–CATALYZED CLEAVAGE OF TRANSMEMBRANE DOMAINS
γ-Secretase is a member of a unique class of proteases that cleave within the transmembrane domain of their membrane-bound substrates. Examples of substrates that undergo intramembrane proteolysis are the sterol regulatory element–binding protein (SREBP), a transcription factor that regulates cholesterol homeostasis, and the Notch-1 protein, which is involved in cell-fate decisions crucial to development (Figure 3⇓) (see 13, ,14). The activity of γ-secretase can be detected by monitoring the production of Aβ40 and/or Aβ42 in cell lines that overexpress either full-length APP or a C-terminal fragment (APP-CTF).
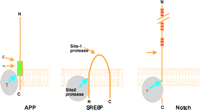
Processing of the APP, SREBP, and Notch. All three integral membrane proteins are proteolytically processed by at least two proteases. Intramembrane cleavage is a unique feature of these proteolytic events. The green box in the APP represents the sequence of Aβ.
Inhibitor studies utilizing difluoroketone-based compounds have indicated that cellular γ-secretase activity is catalyzed by an aspartyl protease (15). It is unclear whether a single protease with dual specificity or two distinct endoproteases generate the C termini of Aβ40 and Aβ42. Although cleavage of APP by β- or α-secretase appears to be a prerequisite for γ-secretase–mediated cleavage (Figure 4⇓), further understanding of γ-secretase activity has been hampered by the technical difficulties of membrane-associated enzymology.
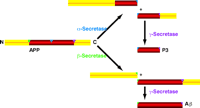
Proteolytic processing of APP. Two pathways of APP processing have been established. APP is cleaved at the extracellular site by α- or β-secretase followed by γ-secretase to release P3 or Aβ peptides, respectively. Thus, inhibition of β- and /or γ-secretase (and activation of α-secretase) will reduce the production of Aβ.
Unraveling the intricacies of the enigmatic γ-secretase is a formidable challenge that requires novel biochemical approaches. The establishment of an appropriate assay in which a substrate with a transmembrane domain can be cleaved by γ-secretase is a key step in elaborating the molecular mechanism of the enzyme. Recently, we reported an in vitro assay, utilizing a recombinant protein substrate, in which γ-secretase activity is detectable in detergent-solubilized cellular membranes (16). The solubilized γ-secretase activity recapitulates the native characteristics of the enzyme in whole cells. Both Aβ40 and Aβ42 are faithfully generated, and the ratio of the two products is similar to that seen in vivo. An aspartyl protease transition-state analog (17) inhibits the solubilized γ-secretase and also blocks γ-secretase–mediated cleavage in cellular studies (16). Pepstatin, a classical inhibitor of aspartyl proteases, also inhibits the activity of the solubilized γ-secretase activity. Other cell-free assays of γ-secretase activity have also been reported, using endogenous APP or APP-CTF as a substrate (17–,20). Gel exclusion chromatography reveals that the solubilized γ-secretase activity elutes as a high molecular–weight complex (16). These studies strongly suggest that γ-secretase activity arises from an aspartyl protease within a macromolecular complex.
PRESENILINS AND γ-SECRETASE ACTIVITY
Presenilin-1 (PS1) and presenilin-2 (PS2) are synthesized as single polypeptide chains with eight putative transmembrane domains. Each presenilin polypeptide undergoes endoproteolysis, whereby the N- and C-terminal cleavage products remain associated as heterodimeric integral membrane proteins (21). The genes that encode PS1 and PS2 were identified because of their association with early-onset AD in humans. Autosomal dominant inheritance of mutations in the PS1 gene is the most common cause of familial early-onset AD (22). More than 70 PS1 mutants have been reported. Likewise, mutations in PS2 can also lead to familial early-onset AD (23).
Considerable investigation indicates that the presenilins play an important role in APP processing and are tightly linked to γ-secretase–mediated cleavage. First, mutations of PS1 and PS2 result in increased production of Aβ peptide—especially Aβ42—in transfected cells and transgenic mice (24–,26); the overproduction of Aβ42 suggests a specific effect on γ-secretase–mediated cleavage. Significantly, the longer form of Aβ is also increased in the brain tissue of patients with PS1-linked familial early-onset AD (27). Second, cultured neurons isolated from PS1-deficient mice exhibit reduced rates of γ-secretase–mediated cleavage of APP (28, ,29). Moreover, γ-secretase activity is abolished in PS1 and PS2 double-knockout cells (30, ,31). Finally, the mutagenesis of two transmembrane-situated aspartates in PS1 was recently shown to inactivate γ-secretase activity (32). On the basis of this finding, it was postulated that PS1 is either an intramembrane aspartyl protease with inherent γ-secretase activity or acts as a unique diaspartyl cofactor for γ-secretase. The possibility that PS1/PS2 are γ-secretase(s) has been widely challenged due to the absence of homology between the presenilins and established proteases as well as to a scarcity of biochemical evidence. It has thus been proposed that presenilins serve as chaperones, controlling the access of γ-secretase to the transmembrane domain of APP, as has been observed in the processing of SREBP by its cleavage activating protein (13). Nevertheless, there is compelling evidence that the presenilins, as is discussed below, act catalytically in the cleavage reaction of APP per se.
WHERE IS THE ACTIVE SITE OF γ-SECRETASE?
Evidence that P presenilins Contain the Aactive Ssite
In the absence of any biochemical evidence to prove that presenilins might be directly associated with γ-secretase activity, early studies demonstrated that APP could be coimmuno-precipitated with PS1 and PS2. Using our in vitro γ-secretase assay, we found that the γ-secretase activity coeluted during gel exclusion chromatography with a PS1 heterodimer and—more importantly—that an anti-PS1 antibody immunoprecipitated γ-secretase activity from our solubilized γ-secretase preparation (16). The fact that the immunocaptured PS1 complex is able to catalyze production of both Aβ40 and Aβ42 further suggests that a single activity is responsible for both cleavage events.
Having established that PS1 is part of the γ-secretase macromolecular complex, we turned to mechanism-based active site inhibitors to probe the catalytic machinery. Hydroxyethylene, hydroxyethylamine, and statine, common transition-state isosteres that mimic the tetrahedral intermediates of proteolysis, have been used in the design of effective inhibitors of aspartyl proteases (Figure 5⇓). The hydroxyl group of these inhibitors results in high affinities for the active sites of proteases such as cathepsin D, renin, pepsin, HIV protease, and BACE. The availability of transition-state analog inhibitors has been crucial to the study of aspartyl proteases, because these enzymes do not covalently bind the tetrahedral reaction intermediates of proteolysis, and so the covalent entrapment of substrate-derived adducts at the active site is not feasible.
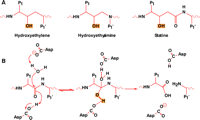
The design of inhibitors of aspartyl proteases.
A. Isosteres that are commonly used to design transition-state analogs of proeolysis. P1 and P1' represent substituents that mimic the amino acid side chains of natural substrates.
B. The reaction mechanism of aspartyl proteases indicating the two aspartates involved in catalysis. An activated water molecule is required for proteolysis. P1 and P1' represent amino acid side chains. The hydroxyl character of the tetrahydryl transitionstate that is mimicked by the isosteres is highlighted.
L-685,458 contains a hydroxyethylene isostere and has been characterized as a transition-state analog that specifically inhibits γ-secretase activity (16, ,17). Photoreactive derivatives of L-685,458 were found to specifically label PS1 and PS2. In the case of PS1, the placement of the photoreactive function within the L-685,458 compound determines which PS1 subunit is modified (Figure 6⇓) (33). Full-length PS1, moreover, does not undergo modification by the photoreactive reagents, whereas PS1 ΔExon9, an enzymically active form of PS1 that lacks the endoproteolytic cleavage site, is readily labeled. These studies thus suggest that full-length PS1 is a zymogen and that a conformational change triggered by endoproteolytic cleavage may result in an active heterodimer whose structure is apparently mimicked by PS1 ΔExon 9. These results further provide compelling biochemical evidence that PS1 contains the active site of γ-secretase; the reactivity of both the C- and N-terminal fragments with photoreactive derivatives of L-685,458, furthermore, indicates that the active site of the protease may be shared between the two subunits of PS1. Similar observations have been made on the basis of other specific inhibitors of γ-secretase (34, ,35).
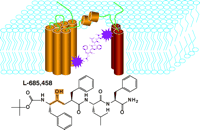
L-685,458 is an apartyl protease transition-state analog that reacts with the presenilins.
Photoreactive groups (shown as pruple star-like geometries) incorportated into L-685,458 result in covalent labeling of PS1, schematized as yellow (N-terminal–derived fragment) and red (C-terminal–derived fragment) subunits. Depending on placement of the photoreactive function, either of the two PS1 subunits can be covalently modified by the inhibitor. The hydroxyethylene isostere of L-685,458 is highlighted (see Figure 5).
Peculiarities of the Presenilins
Because presenilins do not contain the highly conserved signature sequence of Asp-Thr/Ser-Gly-Thr/Ser that typifies aspartyl proteases, some investigators have been reluctant to consider PS1 and PS2 as γ-secretases. Based on the experimental evidence discussed above, however, it seems reasonable to propose that aspartyl proteases may comprise multiple subclasses. Indeed, Steiner and colleagues (36) have recently found that the presenilins share limited homology around one of the active-site aspartates (Gly/Ala-X-Gly-Asp-X-) with bacterial type 4 prepilin peptidase (TFPP), an atypical polytopic membrane-bound aspartyl protease. A second objection to the assignment of γ-secretase activity to the presenilins arose with the observation that the presenilins and the substrates of γ-secretase are not co-localized at the site of enzyme action. This spatial paradox, however, may have been solved by the discovery of small amounts of presenilins on the cell surface (37) and endosomes (38) where γ-secretase acts in vivo. Although genetic, cellular, and biochemical studies strongly suggest that PS1 contains the active site of γ-secretase (Table 1⇓), they do not exclude involvement of other proteins as critical factors of γ-secretase activity. For example, Yu and colleagues (39) have identified a presenilin partner protein named nicastrin that may be involved in γ-secretase activity; however, the exact role of nicastrin in γ-secretase activity remains unknown. It is important, furthermore, to note that the affinity labeling data (see above) cannot rule out the remote possibility that presenilins are merely close in proximity to the active site of γ-secretase. Reconstitution of γ-secretase activity from purified components in vitro may clarify the identity of the proteins that are essential to catalysis, although the definitive elucidation of the γ-secretase reaction mechanism, including identification of catalytic residues, may have to come from the analysis of crystal structures.
Evidence that Presenilins Contain the Active Site of γ-Secretase
Other functions of the presenilins, independent of γ-secretase activity, also deserve consideration; a role in capacitative calcium entry may be one such possibility (see 40). The ability of the presenilins to participate in alternative macromolecular complexes has not been established. γ-Secretase inhibitors, which do not disrupt the physical complex(es) of macromolecules, provide powerful tools to dissect the functions of presenilins. These inhibitors thus offer an advantage over genetic knockout approaches that would eliminate presenilins and their complexes altogether.
γ-SECRETASE, NOTCH, AND AD THERAPY
The Notch signaling pathway plays an essential role in cell fate decisions during embryogenesis, hematopoiesis, and neuronal stem cell differentiation. Presenilins are required for the proteolytic release of the Notch-1 intracellular domain (NICD) from the cell membrane (14). After cleavage, the NICD translocates into the nucleus and acts as a transcription factor. The proteolytic release of NICD is reduced in PS1-deficient cells and abolished in PS1/PS2 double-knockout cells (30, ,31). Reduction of Notch signaling is also observed in presenilin-deficient animal models.
Both APP and Notch proteolysis are blocked by presenilin-binding γ-secretase inhibitors (41). Unfortunately, because both APP and Notch serve as substrates of γ-secretase, inhibition of the protease as an AD therapy may interfere with Notch signaling in adults. Based on the known roles of Notch in adults, the most likely effect would be seen in hematopoiesis. Recent studies have shown that the treatment of fetal thymocyte cultures with γ-secretase inhibitors interferes with T cell development in a manner consistent with the loss or reduction of Notch1 function (42, ,43). Therefore, total inactivation of γ-secretase will most likely be toxic. In addition, it is probable that other γ-secretase substrates will be discovered, which would further complicate the potential for AD therapy. It may nevertheless prove possible to find a therapeutic window in which partial inhibition of γ-secretase activity could reduce Aβ production without obliteration of Notch signaling. Recent reports (40, ,44) that some γ-secretase inhibitors and PS1 mutations have differential effects on Notch and APP processing may reflect the feasibility of developing selective inhibitors of Aβ production. Some of these concerns, as well as the question as to whether interference with the turnover of APP may produce toxic intermediates (45), will be answered in the clinical trials of the γ-secretase inhibitors. Clinical studies of β- and γ-secretase inhibitors should also provide the definitive test of the amyloid hypothesis.
γ-SECRETASE AND REGULATED INTRAMEMBRANE PROTEOLYSIS
While unraveling the proteolytic processing of APP, Notch, and other membrane proteins, a novel signaling paradigm has emerged, namely, regulated intramembrane proteolysis (RIP; Figure 7⇓) (13). RIP requires two proteolytic steps, such that the first step, extracytosolic processing of a polytopic membrane protein, appears to be a prerequisite for the second, intramembrane cleavage. RIP is realized as a mechanism of signal transduction when the cytosolic fragment the intramembrane cleavage is released into the cytoplasm for subsequent translocation into the nucleus to regulate gene transcription. The recent discovery by Cao and Südhof (46) linking the catabolic product of APP cleavage to transcription activation may illuminate the processing of APP as a model of RIP signaling. Currently, two substrates, APP and Notch, have been identified in association with PS-1/γ-secretase cleavage; two other substrates, SREBP and ATF6, are associated with RIP processing by other proteases (13, ,47).
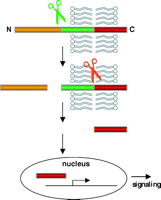
Regulated intramembrane proteolysis (RIP) and signal transduction. Signal transduction by RIP depends on the sequential proteolytic processing of an integral membrane protein. First, an extracytosolic cleavage (green scissors) is implemented, followed by an intramembrane cleavage (red scissors) that releases a cytosolic fragment (shown in red). Upon translocation into the nucleus, this fragment regulates nuclear metabolism, which may culminate in further signaling.
OUTSTANDING ISSUES
Biochemical studies provide compelling evidence that presenilins are novel aspartyl proteases that mediate γ-secretase activity within a macromolecular complex. The identification and characterization of other proteins that associate with the presenilin/γ-secretase complex may help to understand the extraordinary catalysis of APP turnover. In particular, the following questions require investigation: 1) How does water get into the active site of γ-secretase, an apparently hydrophobic environment, to mediate proteolysis? 2) How does γ-secretase recognize the different primary cleavage sequences of APP and Notch? 3) What is the mechanism by which PS1 mutations result in early-onset familial AD, and specifically, in heightened production of Aβ42? 4) Can other proteins in the complex serve as targets for AD therapy? 5) Is γ-secretase a part of the general regulated cellular machinery used to clean up transmembrane fragments? 6) Is it feasible to get a crystal structure of γ-secretase? No doubt, the recognition of γ-secretase as a therapeutic target in AD and as an unusual catalyst will drive these outstanding issues toward resolution.

Yue-Ming Li, Ph.D., is a Senior Research Fellow at Merck Research Laboratories. He received his doctoral degree from the University of California, Berkeley, and pursued postdoctoral training at Harvard Medical School. E-mail yueming_li{at}merck.com; fax 215-652-3082.
Acknowledgments
I thank Drs. Elizabeth Chen Denson, James Bruce, and Ming-tain Lai for discussions and suggestions regarding this manuscript.
- © American Society for Pharmacology and Experimental Theraputics 2001

